Вегетативные клетки бактерий
образованию эндоспор способна только небольшая группа бактерий. Огромное значение эндоспор связано с их термоустойчивостью. В то время как почти все остальные бактерии, а также вегетативные клетки спорообразующих видов гибнут при 80°С (при температуре пастеризации) через 10 мин, терморезистентные эндоспорыпереносят значительно более сильный нагрев; некоторые споры выдерживают даже кипячение на протяжении многих часов. Трудоемкая и дорогостоящая техника стерилизации (обеззараживания) рассчитана на уничтожение эндоспор. С другой стороны, терморезистентность спор предоставляет своеобразную возможность избирательного обогащения культур спорообразующих форм. Землю или иной материал, взятый в определенном месте, на протяжении 10 мин прогревают при 80 или 100°С, что приводит к гибели вегетативных клеток; только терморезистентные споры остаются жизнеспособными и прорастают в подходящей питательной среде.
Классификация бактерий, образующих эндоспоры.Спорообразующие виды, за одним исключением, относятся к палочковидным грам-положительным бактериям. Большинство из них подвижно благодаря перитрихально расположенным жгутикам. Бактерии, принадлежащие к роду Bacillus, — строгие аэробы или факультативные анаэробы. Роды Clostridium и Desulfotomaculum объединяют анаэробные бактерии, способные к образованию спор. Клостридии получают энергию за счет брожения; виды Desulfotomaculum способны получать энергию путем анаэробного дыхания, используя сульфат в качестве акцептора электронов. Sporolactobacillus принадлежит к числу молочнокислых бактерий. Sporosarcina имеет сферические клетки, но по своим физиологическим признакам относится к бациллам. Содержание GC в ДНК клеток, способных образовать споры, удивительно низко; клостридии, у которых ДНК содержит от 22 до 27 мол. % GC, оказываются в конце списка прокариот, расположенных в порядке убывания этого показателя.
Выявление эндоспор.При микроскопическом исследовании споры видны благодаря своему высокому показателю преломления — такому же, как у обезвоженного белка; это указывает на то, что в спорах большое количество богатого белком материала сконцентрировано в малом объеме. Спора содержит почти все сухое вещество материнской клетки, но занимает в 10 раз меньший объем. В сомнительных случаях вопрос о том, имеются ли в клетках истинные эндоспоры, можно решить с помощью специального окрашивания. Если препарат бактерий, фиксированный нагреванием, прокипятить с карболовым раствором фуксина, то споры прочно связывают краситель и не обесцвечиваются даже при обработке этанолом или 1 М уксусной кислотой, тогда как остальное содержимое клетки при этом становится бесцветным.
Спорообразование(споруляция). Споры образуются внутри бактериальной клетки. Этот процесс начинается с накопления белкового материала, поэтому показатель преломления в месте образования споры возрастает. Происходящие при этом метаболические превращения сопровождаются расходованием запасных веществ (поли-3-гидроксимасляной кислоты у аэробов и полисахаридов у анаэробов). В течение первых пяти часов спорообразования значительная часть белков материнской клетки распадается. При этом образуется специфичное для спор вещество — дипиколиновая (пиридин-2,6-дикарбоновая) кислота. В вегетативных клетках эта кислота не встречается. В ходе синтеза дипиколиновой кислоты происходит поглощение ионов кальция; в зрелых спорах эта кислота находится, по-видимому, в виде хелата с кальцием и может составлять 10-15% сухого вещества спор. Дипиколиновая кислота локализована в протопласте споры и имеется только в терморезистентных эндоспорах (рис. 2.45, 2.46).
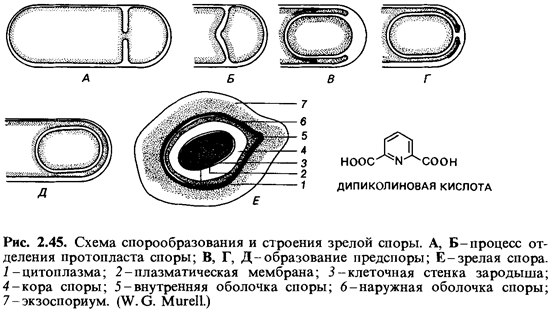
Спорообразование — один из сложнейших процессов дифференцировки бактериальной клетки. Оно начинается с особого неравного деленияклетки (рис. 2.45). В результате впячивания плазматической мембраны часть протопласта отшнуровывается от материнской клетки. Этот протопласт содержит часть ядерного материала — один геном. Образования клеточной стенки между обоими протопластами (как при обычном делении) здесь не происходит. Вместо этого протопласт будущей споры окружается, как бы обрастает, плазматической мембраной материнской клетки. В результате вокруг него оказываются две плазматические мембраны, и каждая из них участвует в синтезе стенки споры. Мембрана протопласта споры синтезирует снаружи от себя стенку зародышевой клетки, а мембрана, происходящая от материнской клетки, синтезирует вовнутрь кору споры (кортекс). Последняя состоит из многослойного пептидогликанового остова, который отличается от каркаса стенок вегетативных клеток помимо прочего степенью сшивки. Наружную оболочку споры образует материнская клетка; эта оболочка в значительной части состоит из полипептидов. Материнская же клетка образует еще один дополнительный тонкий полипептидный покров — экзоспорий; он имеется только у немногих бактерий (например, у Bacillus cereus) и окружает спору в виде свободного чехла. Ввиду такой многослойности покровов не покажется удивительным, что на долю оболочки приходится около половины объема и соответственно сухой массы зрелой споры.
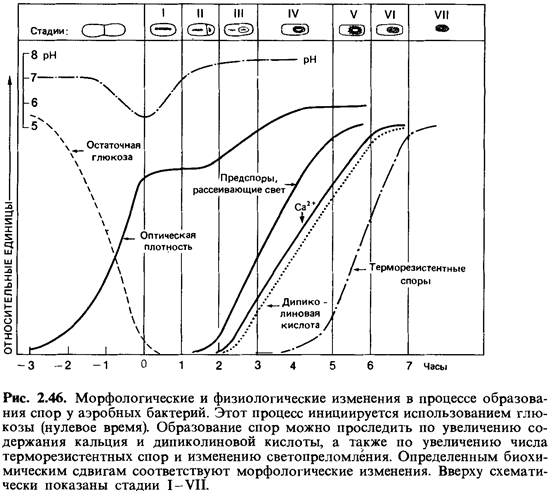
Индукция спорообразования.Споры отнюдь не являются обязательной стадией жизненного цикла бацилл; при благоприятных условиях питания бациллы могут неограниченное время размножаться делением как вегетативные клетки. Образование спор начинается лишь тогда, когда не хватает питательных веществ или когда в избытке накапливаются продукты обмена. Иными словами, оно происходит лишь в тех случаях, когда этому способствуют общие условия. Высыхание не стимулирует споруляцию. Если поместить вегетативные клетки в дистиллированную воду, можно наблюдать «эндотрофную споруляцию», т.е. образование спор за счет накопленных внутри клеток запасных веществ. В таких случаях образование спор, очевидно, вызывается недостатком экзогенного субстрата. Индукция спорообразования осуществляется в течение не скольких часов. Если, например, к суспензии вегетативных клеток Bacillus cereus var. mycoides в первые 5 ч после помещения клеток в воду добавить глюкозу, то образование спор прекратится — добавление субстрата подавляет споруляцию. Глюкоза, добавленная позднее чем через 6 ч, менее действенна: подавление спорообразования не происходит. Индукция (дерепрессия) спорообразования продолжается, и через 10-13 ч после переноса клеток в воду около 90% их образует споры. Следовательно, споруляция регулируется внешними факторами.
Число клеток, образующих споры, во многих случаях возрастает после добавления к среде солей марганца.
Способность образовывать эндоспоры постепенно утрачивается при многократных пересевах вегетативных клеток. Поскольку суспензии спорообразующих микроорганизмов обычно содержат и споры, и вегетативные клетки, перед каждым пересевом культуру, как правило, подвергают кратковременному кипячению. Это способствует сохранению или повышению способности клеток образовывать споры.
Свойства зрелых спор.Споры освобождаются при автолизе материнских клеток. Зрелые споры не проявляют никакой метаболической активности. Они чрезвычайно устойчивы к воздействию высокой температуры, разного рода излучений и химических агентов. Терморезистентность обусловлена очень низким содержанием воды. В спорах Bacillus megaterium всего лишь около 15% воды, т.е. примерно столько же, сколько в шерсти или сухом казеине. Лиофилизированные вегетативные клетки бактерий тоже очень термостойки. Терморезистентность спор приблизительно пропорциональна содержанию в них дипиколиновой кислоты.
Радиорезистентность спор также выше, чем у вегетативных клеток. Она примерно пропорциональна содержанию дисульфидных групп в наружном слое белка. Оболочка споры содержит главным образом белок, богатый цистеином и напоминающий кератин. Химическая же устойчивость эндоспор обусловлена непроницаемостью их оболочки для многих веществ.
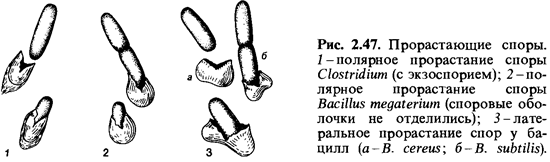
Прорастание спор.Вподходящих средах большинство спор прорастает. Соответствующая предварительная обработка, определенные условия хранения и прогрев могут повысить «всхожесть» спор — увеличить процент прорастания. В случае Bacillus subtilis оптимальными условия ми для стимуляции прорастания спор считают семидневный период покоя и пятиминутный прогрев в воде при 60°С. Другие споры могут быть активированы кратковременным кипячением (10 мин при 100°С). Обработка тепловым шоком должна проводиться непосредственно перед высевом спор, так как процесс активации обратим. Прорастанию спор предшествует поглощение ими воды и набухание. Для прорастания активированных спор в ряде случаев необходимо присутствие глюкозы, аминокислот, нуклеозидов или других соединений. В ходе прорастания спор происходят глубокие физиологические изменения: дыхание и ферментативная активность быстро возрастают; начинается выделение аминокислот, дипиколиновой кислоты и пептидов. При прорастании спор потеря сухого вещества достигает 25-30%. Во время прорастания споры теряют свою термоустойчивость. Выходящая из споры ростовая трубка бывает окружена очень тонкой и, видимо, неполностью сформированной клеточной стенкой, так что в протопласт может, например, проникать даже ДНК. Ростовая трубка может образоваться как в полярном, так и латеральном положении; в одних случаях оболочка споры при этом разрывается, в других ростовая трубка прокалывает ее (рис. 2.47).
Продолжительность жизни спор.Бактерии в виде спор могут дли тельное время находиться в состоянии анабиоза. В земле, которая прилипла к растениям из гербария Кью Гарденс (Англия) и пролежала вместе с ними в сухом состоянии от 200 до 320 лет, было обнаружено лишь небольшое количество жизнеспособных спор Bacillus subtilis и В. liche-niformis. В пробах почвы, хранившихся от 50 до 100 лет, были также обнаружены споры В. coagulans и В. circulans. По данным такого рода экспериментов, в сухой почве за 50 лет хранения до 90% спор теряет жизнеспособность. Судя по этому, одна тонна сухой почвы и через 1000 лет все еще будет содержать жизнеспособные споры.
В сухом состоянии многие бактерии (если не большинство из них) на протяжении ряда лет сохраняют жизнеспособность. Для консервациибактерий в коллекциях вегетативные клетки подвергают, как правило, лиофильной сушке и хранят при комнатной температуре или при низких температурах в вакууме. Как рассчитал еще Беккерель, микроорганизмы при температуре, близкой к абсолютному нулю, могут оставаться жизнеспособными на протяжении миллионов лет. Кратковременные опыты с жидким азотом и экстраполяция их результатов позволяют заключить, что такие предположения вполне оправданны. Бактерии, не переносящие лиофилизации, выдерживают многолетнее хранение в суспензиях при температуре жидкого азота.
Другие покоящиеся формы (цисты, экзоспоры, миксоспоры).Эндоспоры являются длительно переживающими формами бактерий, устойчивыми к высокой температуре, высыханию, облучению и химическим воздействиям. Помимо эндоспор у некоторых других бактерий существуют иные покоящиеся формы — экзоспоры и цисты. Образование экзоспор до сих пор наблюдалось только у метаболизирующих метан бактерий Methylosinus trichosporium. Экзоспоры возникают путем почкования материнской клетки; они сходны по своим свойствам с эндоспорами бацилл. Некоторые бактерии образуют шарообразные толстостенные клетки, называемые цистами. При истощении пищевых ресурсов в цисту превращается вся палочковидная вегетативная клетка, а не только часть ее, как при образовании эндоспоры. Цисты видов Azotobacter, а также Methylocystis устойчивы к высушиванию, механическим нагрузкам и облучению, но не к действию высоких температур. Подобного рода превращение всей клетки в цисту лежит и в основе образования миксоспор из палочковидных вегетативных клеток Myxococcus и Sporocytophaga.
Клетки Arthrobacter (A. globiformis) плеоморфны. При обилии субстрата они растут в виде палочек; когда же субстрат иссякает, появляются кокковидные клетки. Arthrobacter относится к тем бактериям, которые способны некоторое время сохраняться в высохшей почве, пре бывая в состоянии покоя; структурной дифференцировки в данном случае не выявлено.
Споровые покровы в основном состоят из белков и в небольшом количестве из липидов и гликолипидов. Белки покровов обладают высокой устойчивостью к неблагоприятным условиям и обеспечивают спорам защиту от действия литических ферментов, других повреждающих факторов, а также предохраняют спору от преждевременного прорастания. Оказалось, что споры мутантов, лишенные покровов, прорастают сразу же после выхода из материнской клетки, даже если условия для последующего роста неблагоприятны. Кортекс построен в основном из молекул особого типа пептидогликана.
Основное функциональное отличие спор у бактерий и грибов: у бактерий споры обеспечивают переживание в неблагоприятных условиях окружающей среды, у грибов образование спор — способ размножения Микробиология: Учебник дл вузов / О.Д.Сидоренко, Е.Г.Борисенко, А.А.Ванькова, Л.И.Войнова. — М.: Инфа_М, 2005. — с 55.

Рис. 2 — Схема строения зрелой бактериальной эндоспоры
Споры бывают круглыми, овальными или эллиптическими; некоторые снабжены «рёбрами жёсткости», усиливающими устойчивость к механическим воздействиям. При микроскопическом исследовании споры выделяются высоким коэффициентом светопреломления, аналогичный таковому у обезвоженного белка.
В зрелой споре различимы: центральный, плохо окрашиваемый участок (спороплазма), двухслойная ЦПМ и оболочка споры.
Спороплазма (протопласт споры) включает цитоплазму, бактериальную хромосому, системы белкового синтеза и некоторые другие (например, анаэробного энергообразования) Асонов Н.Р. Микробиология: Учебник -4-е изд., перераб. и доп.- М.: КолосC, 2002. — с 65.
Оболочка споры двухслойная: пространство между слоями заполняют гликопептидные полимеры, сходные с пептидогликанами, образующие сетчатую структуру (кортекс), проявляющую высокую чувствительность к лизоциму. Внутренний слой (стенка споры) образован пептидогликанами, аналогичными таковым вегетирующей клетки. Внешний слой (собственно оболочка) образуют кератиноподобные белковые структуры с низкой проницаемостью.
Экзоспориум. У некоторых бактерий материнская клетка образует экзоспориум — двух-трёхслойное желатинообразное покрытие образованное липопротеинами и углеводами и во многом аналогичное капсуле бактерий. При созревании споры экзоспориум может сохраняться в виде пустого и отстающего от споры «мешка».
У бактерий спора, покрытая особо прочной оболочкой, — средство для переживания не6лагоприятных условий. Грибы образуют споры для размножения.
Грибы размножаются половым и бесполым (вегетативным) способом. При вегетативном размножении образуются специализированные репродуктивные структуры — споры — конидии. Они могут располагаться в специализированных вместилищах — спорангиях (эндоспоры) или отшнуровываться от плодоносящих гиф (экзоспоры). Реже наблюдают образование спор внутри клеток (оидии), являющихся сегментами гиф. Дрожжевые клетки размножаются почкованием, мицелий не образуют. Половое размножение включает взаимодействие специализированных клеток, имеющих существенные различия в морфологии у различных грибов и часто используемых как дифференциально-диагностический признак.
Для большинства видов грибов, имеющих медицинское значение, характерно наличие конидий (или экзоспор), являющихся формами неполового размножения. Их классификация во многом основывается на морфологических формах конидий. Их наиболее частые формы- бластопоры, хламидоспоры, артроспоры, конидиоспоры.
Бластоспоры — простые структуры, которые образуются в результате почкования, с последующим отделением почки от родительской клетки, например у дрожжевых грибов.
Хламидоспоры образуются в результате увеличения гифальных клеток с образованием толстой оболочки, защищающей споры от неблагоприятных условий окружающей среды.
Артроспоры — споры, образующиеся путем фрагментации гиф на отдельные клетки. Они встречаются у дрожжеподобных грибов, возбудителя кокцидиоидоза, тканевых форм дерматофитов в волосе, кожных чешуйках и в ногтях.
Конидиоспоры — зрелые наружные споры, возникающие на дифференцированных конидиофорах (конидионосцах), отличающихся от других нитей мицелия по форме и размерам (у аспергилл, пеницилл) или располагающиеся по бокам и на концах любой ветви мицелия, прикрепляясь к ней непосредственно или тонкой ножкой.
К эндоспорам совершенных грибов относятся спорангиоспоры мукоровых грибов, развивающихся в специальных органах (спорангиях), располагающихся на вершине спорангиеносца. Споры освобождаются при разрыве стенки спорангия.
Эндоспоры обнаруживают также у тканевых форм возбудителей кокцидиоидоза. Они развиваются в круглых образованиях — сферулах, при разрыве стенки зрелой сферулы попадают во внешнюю среду.
Спорообразование — способ сохранения определенных видов бактерий в неблагоприятных условиях среды. Эндоспоры образуются в цитоплазме, представляют собой клетки с низкой метаболической активностью и высокой устойчивостью (резистентностью) к высушиванию, действию химических факторов, высокой температуры и других неблагоприятных факторов окружающей среды. При световой микроскопии часто используют метод выявления спор по Ожешко. Высокая резистентность связана с большим содержанием кальциевой соли дипиколиновой кислоты в оболочке спор. Расположение и размеры спор у различных микроорганизмов отличается, что имеет дифференциально-диагностическое (таксономическое) значение. Основные фазы «жизненного цикла» спор — споруляция (включает подготовительную стадию, стадию предспоры, образования оболочки, созревания и покоя) и прорастание, заканчивающееся образованием вегетативной формы. Процесс спорообразования генетически обусловлен.
Микроорганизмы проявляют разную чувствительность к средствам, применяемым для их уничтожения. Существуют видовые различия в чувствительности, а также различия, зависящие от влажности и рН среды, от возраста вегетативных клеток или спор и т.д. Эффективность различных агентов, применяемых для уничтожения микроорганизмов, характеризуют величиной D10 (время, необходимое для того, чтобы в определенной популяции при определенных условиях среды вызвать гибель 90% клеток).
Полная или частичная стерилизация осуществляется с помощью:
1) влажного жара,
5) различных химических средств.
1. Влажный жар Вегетативные клетки большинства бактерий и грибов гибнут через 5-10 мин уже при температуре около 60°С, споры дрожжей и мицелиальных грибов лишь при температурах выше 80°С, а споры бактерий — выше 120°С (15 мин). Время воздействия влажным жаром, необходимое для уничтожения спор некоторых видов бактерий, отличающихся чрезвычайной термоустойчивостью. При этом следует учитывать, что окончательный результат стерилизации зависит также от степени загрязнения обрабатываемого материала, т.е., например, от числа терморезистентных спор: чем их больше, тем длительнее должен быть нагрев. Для достижения температур выше точки кипения воды пользуются автоклавом. Температура насыщенного пара зависит от давления.
При доступе воздуха определенному давлению соответствует значительно более низкая температура. Продолжительность стерилизации, естественно, зависит от объема (теплоемкости) сосудов, в которых ее проводят.
Тиндализация — нередко удается достичь того же эффекта дробной стерилизацией в текучем паре при 100°С . Жидкость стерилизуется в этом случае при 100°С три дня подряд по 30 мин ежедневно; в промежутках между нагреваниями ее хранят в термостате, для того чтобы споры проросли, а затем вегетативные клетки были уничтожены при следующем нагревании.
Для многих целей довольствуются частичной стерилизацией, т.е. уничтожением вегетативных форм микроорганизмов. Такого эффекта обычно достигают путем пастеризации — выдерживания в течение 5-10 мин при 75 или 80°С.
Пастеризацией частично стерилизуют, в частности, молоко; однако, чтобы не испортить его вкуса, время воздействия в этом случае сокращают. Применяют два метода пастеризации молока: кратковременное нагревание (20 с при 71,5-74°С) и сильное нагревание (2-5 с при 85-87°С). Стерилизации молока добиваются в результате сверхсильного нагревания: При этом в молоко вводят перегретый водяной пар, доводя температуру смеси до 135-150°С. Молоко подвергается действию этой температуры в течение 1 -2 с. Затем, пропуская молоко через форсунку, понижают давление и одновременно охлаждают молоко; при этом из него удаляется вода, введенная в виде пара.
Способы консервирования ягод и косточковых плодов тоже следует рассматривать как частичную стерилизацию. При обычном нагревании консервных банок в течение 20 мин при 80°С гибнут только вегетативные клетки и споры многих грибов, в то время как споры бактерий остаются жизнеспособными. Прорастанию бактериальных спор препятствуют низкие значения рН, обусловленные присутствием кислот во фруктовом соке. На пастеризованной клубнике часто появляется так называемый «клубничный гриб» Byssochlamys nivea. Его аскоспоры выдерживают 86°С; при этой температуре составляет 14 мин.
2. Сухой жар. При стерилизации сухим жаром бактериальные споры переносят более высокие температуры и притом дольше, чем при стерилизации влажным жаром. Поэтому жаростойкую стеклянную посуду, порошки, масла и т. п. стерилизуют в течение 2 ч при 160°С в сухом стерилизаторе. В тех случаях, когда это позволяет стерилизуемый материал, в настоящее время применяют 30-минутный нагрев при 180°С. Как показывает опыт, при этом погибают все споры. Стерилизация жаром основана на коагуляции клеточных белков.
3. Фильтрация. Растворы, содержащие термолабильные вещества, удобнее всего стерилизовать фильтрованием. В лабораториях и для стерилизации питьевой воды используют фильтры Беркефельда (из прессованного кизельгура). Часто употребляют также асбестовые пластинки (в фильтрах Зейца), стеклянные фильтры и мембранные фильтры. Некоторые из них выпускаются с различной величиной пор, что позволяет даже разделять организмы разной величины и формы.
4. Облучение. Для полной или частичной стерилизации применяют ультрафиолетовые, рентгеновские и гамма-лучи. В лабораторных условиях наибольшее значение имеют ультрафиолетовые лучи. В спектре УФ-ламп преобладает излучение в области 260 нм, поглощаемое главным образом нуклеиновыми кислотами и при достаточно длительном воздействии вызывающее гибель всех бактерий. УФ-облучение используется для частичной стерилизации помещений; при этом бактерии погибают очень быстро, а споры грибов, гораздо менее чувствительные к ультрафиолету, значительно медленнее. Ионизирующее излучение применяют для стерилизации пищевых продуктов и других компактных материалов.
5. Химические средства. При стерилизации пищевых продуктов, лекарственных препаратов и разного рода приборов, а также в лабораторной практике оправдало себя применение окиси этилена, которая убивает и вегетативные клетки, и споры, но действует только в том случае, если подвергаемые стерилизации материалы содержат некоторое количество (5-15%) воды. Окись этилена применяют в виде газовой смеси (с N2 или СО2), в которой ее доля составляет от 2 до 50%.
Для сохранения термолабильных веществ, содержащихся в питательных средах, в практику была введена стерилизация пропиолактоном. Он значительно активнее окиси этилена, но обладает, видимо, довольно сильным канцерогенным действием и вызывает ряд других побочных физиологических эффектов. Его добавляют в количестве 0,2% в готовые питательные среды, которые затем инкубируют 2 ч при 37°С. Если оставить среду на ночь, пропиолактон полностью разложится.
Углеводы при этом не затрагиваются. Напитки стерилизуют также дизтилпирокарбонатом (0,003-0,02%) Асонов Н.Р. Микробиология: Учебник -4-е изд., перераб. и доп.- М.: КолосC, 2002. — с 44.
Для стерилизации семян, используемых при выращивании стерильных растений, пригодны такие обычные антимикробные средства, как бромная вода (1%), сулема (HgCl2; 1%-ный раствор в спирте), AgNO3 (0,05%), гипохлорит кальция [1% Са(С1О)2], успулун и др., которыми воздействуют в течение 5-30 мин. Перед этим семена следует обработать мылом или другим поверхностно-активным веществом, чтобы обеспечить полное смачивание поверхности Гусев М.В. Микробиология: Учебник для вузов. — 4-е изд., — М.: Академия, 2003. — с 29.
Читайте также:
- Эндоспоры и другие покоящиеся формы бактерий. Процессы образования и формирования эндоспор. Значение спор у бактерий.
Строение. Эндоспора —метаболически неактивная(анабиотическая, покоящаяся) форма бактерий. Эндоспоры образуются при попадании вегетативной клетки в неблагоприятную внешнюю среду.
Факторы, стимулирующие спорообразование:
– недостаток питательных веществ (прежде всего углерода и азота);
– накопление в среде в большом количестве токсичных продуктов обмена веществ бактерий;
– минеральные соли, содержащие ионы кальция, магния, калия, марганца, железа, цинка, меди;
– минимальные концентрации некоторых редких металлов (кобальта, лития, кадмия, никеля);
– парааминобензойная и фолиевая кислоты.
Способностью к спорообразованию обладает небольшое число как патогенных бактерий (B. anthracis, C. perfringens, C. tetani, C. botulinum), так и непатогенных бактерий (сапрофитные представители рода Bacillus и некоторые кокки) (рис. 30). Спорообразование — непостоянный признак: у бацилл сибирской язвы споры образуются при 12–42 0 С, эта способность теряется после длительного выращивания при 42,5 0 С.

Рис. 30. Виды эндопор:
1 – центральная спора, вегетативная клетка не лизируется, 2 – терминальная спора, 3 – «теннисная ракетка», 4 – центральная спора, вегетативная клетка лизируется, 5 – «барабанная палочка», 6 – свободные споры
Споры очень резистентны к физическим (высушиванию, высокой температуре, радиации, вакууму) и химическим (дезинфектантам, токсическим веществам, кислороду) факторам. Это позволяет им выжить в условиях, губительно действующих на вегетативные клетки.
Терморезистентность эндоспор обусловлена:
– практически полным отсутствием свободной воды;
– большим содержанием кальциевой соли дипиколиновой кислоты, которая не встречается у вегетативных клеток;
– особым строением белка;
– особым строением пептидогликана кортекса;
– многослойностью и толщиной оболочки.
Эндоспоры образуются внутри бактериальной клетки. Образование эндоспор у бактерий не является способом размножения. Как правило, внутри бактериальной клетки образуется только одна спора. У отдельных видов рода Clostridium обнаружены клетки с двумя и более спорами.
Объем споры в 10 раз меньше, чем вегетативной клетки. Структурно спора представляет ДНК, окруженную многослойной оболочкой (рис. 31).

Рис. 31. Схематическое изображение строения споры:1 — экзоспориум; 2 — слои споровой оболочки; 3 — внешняя мембрана споры; 4 — кортекс; 5 — внутренняя мембрана споры; 6 — сердцевина
Строение сердцевины спор сходно у разных видов. Объем оболочки споры достигает 50% от всего объема споры. Строение оболочек спор у разных видов сильно различается. Оболочка определяет форму споры, защищает ее от действия литических ферментов и от преждевременного прорастания. Споры мутантных штаммов, лишенные оболочек, прорастают даже в неблагоприятной для роста среде, но проросшие клетки в такой среде погибают.
Споруляция — процесс интенсивного распада белков материнской клетки с одновременным образованием новых белковых компонентов. Поэтому по антигенной структуре споры и вегетативные клетки одного и того же вида различны. Имеются также четкие различия между спорами разных видов. Процесс спорообразования генетически обусловлен. Он длится 18–20 часов, внешние источники питания и энергии не требуются.
Стадии спорообразования (споруляции):
1.Подготовительная. Перед спорообразованием резко снижается синтез ДНК и деление клеток прекращается. Происходит накопление большого количества запасного питательного вещества — гранулезы, откладывающегося в виде гранул. ЦП становится зернистой. Клетки резко увеличиваются в размерах, принимают вид лимона либо барабанной палочки, иногда клетки сохраняют обычную палочковидную форму.
| У спорообразующей бактерии образуется дополнительный нуклеоид, который отходит к одному из полюсов клетки (рис. 32). |
 |
| Рис. 32. Образование дополнительного нуклеоида |
Вокруг дополнительного нуклеоида в ЦП спорообразующей клетки образуется спорогенная зона — уплотненный участок ЦП, не имеющий свободной воды.
| 2. Стадия предспоры (проспоры). В ЦПМ, ближе к одному из полюсов клетки, возникает инвагинация. В этом процессе участвуют мезосомы, которые спаивают сближающиеся участки инвагинированных мембран и образуют споровую перегородку (септу), разделяющую клетку на два протопласта, каждый из которых содержит одну хромосому (рис. 33). |
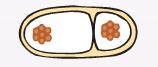 |
| Рис. 33.Отделение протопласта |
Затем материнская клетка поглощает отсеченный участок ЦП с ядром (рис. 34). Это осуществляется путем роста и продвижения периферических участков мембраны материнской клетки по направлению к полюсу клетки. Эта стадия спорообразования обратима.
| Если после образования септы добавить к спорулирующей культуре антибиотик левомицетин, то синтез белков и рост мембраны будет подавлен. Движение периферических участков мембраны, поглощающей отсеченный участок протопласта, будет остановлено. |
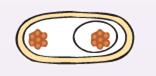 |
| Рис. 34. Материнская клетка поглощает протопласт |
В результате начавшийся процесс спорообразования превратится в обычный процесс вегетативного деления клеток.
| В конце этой стадии сближающиеся участки мембраны сливаются и меньший из протопластов (проспора) покрывается второй оболочкой, синтезируемой мембраной материнской клетки (рис. 35). |
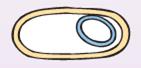 |
| Рис. 35. Проспора покрывается второй оболочкой |
При этом спорогенная зона изолируется от остальной части ЦП. Так внутри клетки возникает новая клетка-проспора, окруженная, в отличие от материнской, двумя трехслойными мембранами — внутренней и внешней. У одних видов проспора остается у полюса клетки, у других видов занимает центральное положение. С этого момента начинается необратимая фаза споруляции.
| 3. Образование кортекса— оболочки между внутренней и внешней мембранами проспоры, состоящей из особых молекул пептидогликана (рис. 36). Кортекс играет защитную роль: он предохраняет сердцевину спор от литических ферментов. |
 |
| Рис. 36. Образование кортекса |
4. Стадия созревания.С внешней стороны наружной мембраны споры образуется внешняя плотная споровая оболочка, в состав которой входят белки, липиды и кальциевая соль дипиколиновой кислоты.
| Снаружи у многих видов анаэробов спора покрывается еще одной дополнительной полипептидной оболочкой — экзоспориумом (рис. 37). |
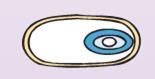 |
| Рис. 37. Образование экзоспориума |
Он имеет вид многослойного чехла, в котором расположена спора. Возникает экзоспориум на ранней стадии формирования споры в виде небольшого пузырька на внешней мембране проспоры, затем пузырек разрастается и охватывает спору со всех сторон. После механического удаления экзоспориума процесс прорастания спор не нарушается.
| 5. Отмирание вегетативной части клетки.На завершающей стадии спорообразования происходит резкое возрастание активности литических ферментов, полностью разрушающих материнскую вегетативную клетку (рис. 38). |
 |
| Рис.38. Свободная спора в стадии покоя |
Спора выходит в окружающую среду, где может сохраняться потенциально жизнеспособной тысячи лет (стадия покоя).
Генетический контроль спорообразования. Синтез новых белковых веществ, споровых оболочек, кортекса и экзоспориума осуществляется при активном контроле генома материнской клетки. Роль материнской клетки сводится к тому, чтобы отделить от себя дочернюю клетку, оказавшуюся внутри ее ЦП. Но так как удалить проспору за пределы клеточных границ через ЦПМ и КС стенку невозможно, то материнская клетка изолирует дочернюю от себя, окружая проспору мощными слоями белковой оболочки и переводя ее в анабиотическое состояние. Но это не спасает материнскую клетку от гибели. Хотя она продолжает еще длительное время активно функционировать, даже иногда способна образовать еще одну спору, но все же она не в состоянии вернуться к вегетативному развитию.
Прорастание споры.При попадании в благоприятные условия спора начинает прорастатьв вегетативную клетку. Прорастание длится 4–5 часов, т. е. в 5 раз быстрее, чем спорообразование. Обычно не все споры, вносимые в благоприятную среду, прорастают сразу. Некоторые споры остаются непроросшими в течение различного времени, не теряя своей жизнеспособности.
Способ и время прорастания молодых спор являются постоянными и могут служить видовыми признаками. Если перенести только сформировавшиеся споры в благоприятную питательную среду, их прорастание начинается не сразу, а через 2–3 часа. Старые споры прорастают более длительно.
Стадии прорастания споры:
1. Набухание, связанное с увеличением в споре количества воды.
2. Дерепрессия генома и активация ферментов, участвующих в энергетическом и пластическом метаболизме. На начальной стадии прорастания спор происходит выделение в среду экссудата прорастания. Половину его сухой массы составляет дипиколинат кальция, остальная часть экссудата прорастания представлена свободными аминокислотами, высокомолекулярными и низкомолекулярными белковыми веществами.
Активация ферментов происходит при обработке спор незначительными количествами некоторых веществ (L–аланина, пуриновых нуклеозидов, глюкозы, аденозина), кипячением или пастеризацией, умеренным воздействием ультразвука. Под действием активаторов дипиколиновая кислота, связывающая ферменты, подвергается деполимеризации и неактивная ферментная система превращается в активную.
Напротив, наличие в среде некоторых веществ (мышьяковистых соединений, углекислого натрия, высоких концентраций ряда аминокислот — глицина, метионина, цистеина и валина, D–аланина) угнетает процесс прорастания спор. Механизм угнетающего действия ряда факторов на прорастание спор окончательно неясен.
3. Разрушение плотных оболочек споры под влиянием давления, вызванного ростом. Через образовавшееся отверстие в оболочке споры молодая клетка (проросток, ростовая трубка) выходит в среду и начинает расти (рис. 39). При этом из одной споры образуется одна вегетативная клетка,которая начинает делится (рис. 40).
 |
 |
| Рис. 39. Проросток споры | Рис. 40. Юная вегетативная клетка |
Выросты на спорах. Особенностью спорообразования у анаэробов является формирование на спорах выростов разнообразной формы в области контактирующих мембран материнской клетки и проспоры. Каждому виду анаэробных бактерий свойствен свой тип строения выростов, этот признак наследственно закреплен и очень устойчив. Выросты состоят в основном из белка, в веществе выростов также обнаружены полисахариды и липиды.
Предположения о функции выростов на спорах:
– ответственны за контакт клеток в системе спорангий — спора,
– подают споре при благоприятных условиях «команду» на прорастание,
– играют важную роль в созревании спор, участвуют в формировании споровых покровов и кортекса,
– являются результатом нарушений в нормальном обмене веществ.
Колпачки на спорах. На спорах анаэробов образуются колпачки ячеистого строения. Ячейки — мешочки, заполненные газообразным веществом. Функция колпачков — увеличение подвижности и плавучести спор в водной среде, что способствовует их распространению в природе. Подобно понтонам ячеистые колпачки не позволяют спорам тонуть. Споры с колпачками не удается осадить даже при центрифугировании.
Функции эндоспор:
1. Способность бактерий образовывать споры позволяет сохранить наследственную информациюпри попадании вегетативной клетки в неблагоприятные условия внешней среды и способствует сохранению вида. Споры могут оставаться жизнеспособными в течение десятков, сотен и даже тысяч лет. Жизнеспособные споры были выделены из египетских мумий, трупов мамонтов.
2. Длительное существование спор бактерий в покоящемся состоянии в окружающей среде имеет существенное эпидемиологическое значение. Попадание спор патогенных бактерий в глубокие колотые и резанные раны, в которых создается анаэробиоз, может привести к возникновению анаэробной раневой инфекции (столбняка или газовой гангрены).
3. Способность бактерий образовывать споры, различающиеся по форме, размерам и локализации в клетке, является таксономическим видовым признаком, который используется для идентификации.
Форма спор может быть овальной, шаровидной, яйцевидной, цилиндрической.
Размер спор у бацилл приблизительно равен диаметру клетки, поэтому клетка не меняет формы. У клостридий размер спор превышает диаметр клетки, поэтому клетка расширяется в середине и принимает форму веретена.
Расположение спор в клетке может быть центральное (у сибиреязвенной бациллы), терминальное (у клостридий столбняка), субтерминальное (у клостридий ботулизма и газовой гангрены).
Выявление эндоспор:
1. При обычных способах окраски споры не прокрашиваются и имеют вид неокрашенных пустот внутри окрасившихся вегетативных клеток, так как плотная оболочка спор непроницаема для воды. Споры видны благодаря своему высокому показателю преломления — такому же как у обезвоженного белка. Это указывает на содержание в спорах бактерий большого количества богатого белком материала, сконцентрированного в малом объеме. Спора содержит почти все сухое вещество материнской клетки, но занимает в 10 раз меньший объем.
2. Специальные методы окраски используют в сомнительных случаях. При этом используют протравы, которые разрыхляют оболочку споры и облегчают проникновение красителя. Окрасившиеся споры обладают кислотоустойчивостью в отличие от вегетативного тела микробной клетки, обесцечивающегося под действием кислоты.
Окраска по Ожешко:
| – на высушенный нефиксированный мазок (толстый, на краю стекла) наносят несколько капель 0,5 % HCl и подогревают 1–2 мин до закипания, остатки кислоты сливают; – остывший препарат промывают водой, подсушивают, фиксируют на пламени горелки; далее красят как по Цилю-Нильсену, для докрашивания можно использовать 1 % малахитовый зеленый. Окрасившиеся споры (рубиново-красные) обладают кислотоустойчивостью в отличие от вегетативных тел микробных клеток (синих или зеленых) (рис.41). |
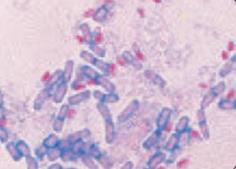 |
| Рис. 41. Окраска по Ожешко |
3. Фазово-контрастная микроскопия.
4. Электронная микроскопия.
Дата добавления: 2015-04-25 ; Просмотров: 3330 ; Нарушение авторских прав? ;
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет

